



Myelodysplastic syndromes complicated by pancreatic iron overload: a case report and review of the literature
-
摘要: 胰腺铁过载是一种罕见的疾病,以糖尿病首发,伴有胰腺形态学改变。临床上因缺乏对胰腺铁过载的认识和诊断标准,往往导致误诊,从而带来不必要的经济、身体和精神负担。本研究报告1例临床上被误诊为胰腺癌的骨髓增生异常综合征并发胰腺铁过载病例,旨在提高疾病认识和降低误诊率。Abstract: Pancreatic iron overload is a rare disease, presented with diabetes as the first symptom and with altered pancreatic morphology. Clinically, the lack of awareness and diagnostic criteria for pancreatic iron overload often leads to misdiagnosis of pancreatic disease, as well as unnecessary economic, physical, and mental burden. Here, we report a case of myelodysplastic syndromes complicated by pancreatic iron overload that was clinically misdiagnosed as pancreatic cancer, aiming at raising awareness and reducing the rate of misdiagnosis.
-
Key words:
- pancreatic cancer /
- myelodysplastic syndromes /
- iron overload
-

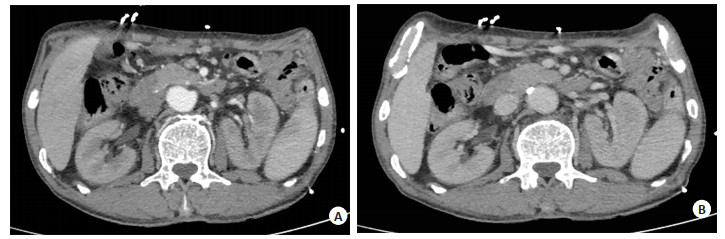
图 1 腹部增强CT图
A: 胰颈部局部密度欠均匀, 增强扫描动脉期胰颈部见团块状弱强化区, CT值约65 Hu; B: 门脉期及静脉期病灶呈轻度渐进性延迟强化.
Figure 1. The abdominal enhanced CT.

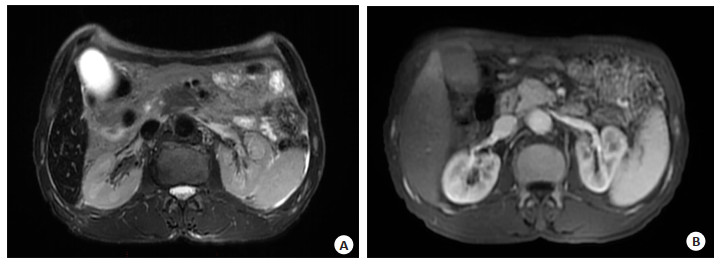
图 2 腹部增强MRI
A: 胰腺颈体似见约29 mm×24 mm稍长T1稍长T2信号影, 增强扫描呈渐进中度强化, 动态增强扫描动脉期低于正常胰腺实质强化; B:肝脏于T2WI像呈弥漫低信号, 同相位信号弥漫低于反相位.
Figure 2. The abdominal MRI.
-
[1] Martín M, Millan A, Ferraro F, et al. Leukocyte telomere length is associated with iron overload in male adults with hereditary hemochromatosis[J]. Biosci Rep, 2020, 40(10): BSR20201916. doi: 10.1042/BSR20201916 [2] Zhang L, Hou L, Liu Z, et al. A mitophagic response to iron overload-induced oxidative damage associated with the PINK1/Parkin pathway in pancreatic beta cells[J]. J Trace Elem Med Biol, 2020, 60: 126493. doi: 10.1016/j.jtemb.2020.126493 [3] Bahr TM, Christensen RD, Ward DM, et al. Ferritin in serum and urine: a pilot study[J]. Blood Cells Mol Dis, 2019, 76: 59-62. doi: 10.1016/j.bcmd.2019.02.001 [4] Pepe A, Pistoia L, Gamberini MR, et al. The close link of pancreatic iron with glucose metabolism and with cardiac complications in thalassemia major: a large, multicenter observational study[J]. Diabetes Care, 2020, 43(11): 2830-9. doi: 10.2337/dc20-0908 [5] Healy GM, Kannengiesser SAR, Espin-Garcia O, et al. Comparison of Inline R2* MRI versus FerriScan for liver iron quantification in patients on chelation therapy for iron overload: preliminary results[J]. Eur Radiol, 2021, 31(12): 9296-305. doi: 10.1007/s00330-021-08019-0 [6] Bas M, Gumruk F, Gonc N, et al. Biochemical markers of glucose metabolism may be used to estimate the degree and progression of iron overload in the liver and pancreas of patients with β-thalassemia major[J]. Ann Hematol, 2015, 94(7): 1099-104. doi: 10.1007/s00277-015-2342-y [7] Lunova M, Schwarz P, Nuraldeen R, et al. Hepcidin knockout mice spontaneously develop chronic pancreatitis owing to cytoplasmic iron overload in acinar cells[J]. J Pathol, 2017, 241(1): 104-14. doi: 10.1002/path.4822 [8] Frazer DM, Wilkins SJ, Darshan D, et al. Stimulated erythropoiesis with secondary iron loading leads to a decrease in hepcidin despite an increase in bone morphogenetic protein 6 expression[J]. Br J Haematol, 2012, 157(5): 615-26. doi: 10.1111/j.1365-2141.2012.09104.x [9] Wang XY, Zhang MZ, Woloshun RR, et al. Oral administration of ginger-derived lipid nanoparticles and Dmt1 siRNA potentiates the effect of dietary iron restriction and mitigates pre-existing iron overload in Hamp KO mice[J]. Nutrients, 2021, 13(5): 1686. doi: 10.3390/nu13051686 [10] Piolatto A, Berchialla P, Allegra S, et al. Pharmacological and clinical evaluation of deferasirox formulations for treatment tailoring[J]. Sci Rep, 2021, 11(1): 12581. doi: 10.1038/s41598-021-91983-w [11] Vadolas J, Ng GZ, Kysenius K, et al. SLN124, a GalNac-siRNA targeting transmembrane serine protease 6, in combination with deferiprone therapy reduces ineffective erythropoiesis and hepatic iron-overload in a mouse model of β-thalassaemia[J]. Br J Haematol, 2021, 194(1): 200-10. doi: 10.1111/bjh.17428 [12] Takpradit C, Viprakasit V, Narkbunnam N, et al. Using of deferasirox and deferoxamine in refractory iron overload thalassemia[J]. Pediatr Int, 2021, 63(4): 404-9. doi: 10.1111/ped.14444 [13] Dai EY, Han L, Liu J, et al. Ferroptotic damage promotes pancreatic tumorigenesis through a TMEM173/STING-dependent DNA sensor pathway[J]. Nat Commun, 2020, 11(1): 6339. doi: 10.1038/s41467-020-20154-8 -
 下载:
下载:
 点击查看大图
点击查看大图
图(3)
计量
- 文章访问数: 145
- HTML全文浏览量: 122
- PDF下载量: 8
- 被引次数: 0